
At depths where crushing pressure renders sunlight a distant memory, a sulfide buttress rises from fractured basalt along a mid-ocean ridge spreading center, its surfaces alive with hundreds of yeti crabs — *Kiwa* species — packed flank to flank in the warmest corridors of vent effluent. Their elongated setae, dense with episymbiotic bacteria, sweep continuously through mineral-laden water, a behavior that amounts to agriculture in the dark: cultivating chemolithotrophic microbes that oxidize hydrogen sulfide and fix carbon where no photon has penetrated for millions of years. The rock itself is layered in silver-white bacterial mats and pale mineral crusts — anhydrite, pyrite, barite — their crystalline surfaces softly luminous beneath the ember-orange chemiluminescence bleeding from nearby black smoker columns, where superheated fluid at several hundred degrees Celsius meets near-freezing abyssal water and precipitates a perpetual dark storm of metal sulfides. Scattered cyan-green bioluminescent pinpoints from peripheral fauna punctuate the gloom beyond the vent field, where marine snow and fine hydrothermal particulates drift in absolute silence through water pressing inward at three hundred atmospheres or more. This community exists entirely on chemical energy drawn from Earth's interior — a self-contained biosphere unconnected to the sun, older in its metabolic logic than almost any ecosystem we know, and complete in itself long before anything observed it.

Where sunlight has never reached and pressure exceeds three hundred atmospheres, a vast shallow depression in the volcanic seafloor lies carpeted wall to wall with thousands of white clams — Calyptogena and their kin — their chalky shells packed so densely they form a living mosaic across dark basalt and sulfide crust. These bivalves owe nothing to photosynthesis; endosymbiotic bacteria housed within their gill tissue harvest hydrogen sulfide seeping upward through the sediment, converting chemical energy directly into organic matter in a process of chemosynthesis that has sustained vent communities since long before complex animal life diversified. Above the clam bed, mineral-rich fluid rises in wavering, refractive curtains, distorting the water column in slow shimmering veils and carrying faint milky hazes of microbial film and precipitating minerals at the point where superheated chemistry meets cold abyssal seawater. Sparse bioluminescent particles drift freely through the black-blue water — brief sparks of cyan and green tracing invisible currents — while marine snow settles in imperceptible suspension all around, and a dim chemiluminescent warmth bleeds upward from the vented ground itself. In the far distance, the silhouettes of black smoker chimneys rise against impenetrable darkness, their plumes dissolving silently into a world that needs nothing from above to persist, reproduce, and thrive.

At depths where pressure exceeds 250 atmospheres and the last photon of sunlight faded kilometers above, the seafloor fractures open along a mid-ocean ridge, and the planet breathes. Towering sulfide chimneys — some rising ten meters or more from the basalt — exhale superheated fluid at 350°C, a temperature that would flash to steam at the surface but here remains liquid under crushing hydrostatic force, the black plumes billowing upward in roiling convection columns through water of near-freezing stillness. Along the edges of the most turbulent eddies, faint copper-blue chemiluminescence traces chemical oxidation reactions between the mineral-laden fluid and the surrounding seawater, the only light this world has ever known. At the base of the spires, white clams of the genus *Calyptogena* spread across fractured basalt in dense beds, their hemoglobin-rich tissues adapted to bind hydrogen sulfide and ferry it to endosymbiotic bacteria that fix carbon without a single ray of sunlight — a food web sealed entirely within the chemistry of the Earth itself. *Riftia pachyptila* tubeworms anchor in the crevices nearby, and pale yeti crabs, *Kiwa* sp., press their setae-covered claws against warm ledges, farming the chemosynthetic microbial films that coat every surface in this silent, pressurized cathedral the planet built for itself, long before anything above the waves existed to witness it.

Beyond the main ventfield, where the last traces of chemosynthetic warmth fade into basaltic cold, a single anglerfish hangs suspended in the crushing stillness of water pressing down at hundreds of atmospheres — her pale cyan-green esca the only unambiguous light in this world, pulsing with bioluminescent bacteria cultivated within a modified dorsal spine through millions of years of deep evolution. That luminous lure, drifting perhaps two kilometers below the surface where photons from the sun are an irrelevance, faintly catches the wet charcoal of her skin, the glassy indifference of her eye, and the translucent edges of fins adapted to a life of near-perfect motionlessness amid sparse, drifting marine snow. Far beyond her, several black smoker chimneys rise as dark silhouettes against their own faint orange-red chemiluminescent shimmer, superheated plumes at 350 °C precipitating metal sulfides the moment they meet near-freezing ambient seawater, building the towering mineral structures that anchor dense communities of Riftia tube worms, chemosynthetic clams, and yeti crabs — ecosystems sustained entirely by the oxidation of hydrogen sulfide rather than sunlight. Between anglerfish and ventfield, scattered sulfide mounds and ash-dark sediment stretch across fresh pillow basalt on a mid-ocean ridge axis, a volcanic landscape that continuously reshapes itself along the planet's spreading boundaries, indifferent to any witness and luminous only on its own chemical terms.

Along the mid-ocean ridge, a ribbon of freshly erupted basalt lies exposed across the seafloor, its glassy black surface still radiating a dull orange-red thermal glow where rock temperatures have not yet surrendered entirely to the near-freezing abyss — a boundary where 350 °C hydrothermal fluid meets water close to 2 °C, producing violent chemical precipitation and the dense, mineralite plumes of black smokers rising as dark sulfide towers above the new crust. Fine sulfur grains settle like pale dust into the folded folds and fractures of pillow lava, while gossamer microbial films — the foundational chemosynthetic organisms of this ecosystem — coat warm surfaces in a faint iridescent sheen, converting hydrogen sulfide into organic energy in the complete absence of sunlight. Clustered around the thermal gradient, scarlet-plumed Riftia pachyptila tube worms stand in pale mineral tubes, their hemoglobin-rich plumes drawing sulfide-laden water toward the endosymbiotic bacteria living within their trophosome tissues, while white Calyptogena clams lie half-buried in sulfide-rich sediment and shaggy yeti crabs patrol the warm basalt margins, cultivating chemosynthetic bacteria across their setae-covered claws. At pressures exceeding 200 to 400 atmospheres, the water column beyond the vent field dissolves from deep cobalt into absolute black, punctuated only by occasional cyan-green bioluminescent pulses and the slow drift of marine snow and mineral particles through immense, indifferent stillness — a self-sustaining world of volcanic heat and chemical energy that has persisted on this planet for hundreds of millions of years, entirely without witness.

Where tectonic plates pull apart along a mid-ocean ridge, superheated fluid charged with hydrogen sulfide, methane, and dissolved metals erupts through fractured basalt at temperatures approaching 350 °C, building towering sulfide chimneys whose billowing plumes carry a faint ember-glow of chemiluminescence into water held near freezing by pressures exceeding 200 atmospheres. Beside one such outcrop, the skeleton of a great whale has come to rest on the abyssal sediment, its bleached and mineral-stained ribs half-buried in black mud, transformed now into an ecosystem of its own — a whale fall in its sulphophilic stage, where the lipid-rich bones sustain dense chemosynthetic microbial mats that in turn feed hagfish coiling through the vertebral arches, amphipods flickering in pale constellations across the skull, and squat lobsters whose blue-green bioluminescence traces each rib like a living diagram. At the vent base, colonies of *Riftia pachyptila* tube worms extend their scarlet plumes into the sulfide-laden gradient, flanked by white vesicomyid clams and pale Yeti crabs combing bacterial films from the chimney flanks — all organisms whose metabolisms are coupled not to sunlight but to the chemical energy bleeding from the planet's interior. Fine mineral particles and marine snow drift through absolute darkness beyond these intimate pools of cold light, and the scene unfolds without witness, existing entirely within itself, a convergence of geological heat and biological patience at a depth where the ocean has never required an audience.
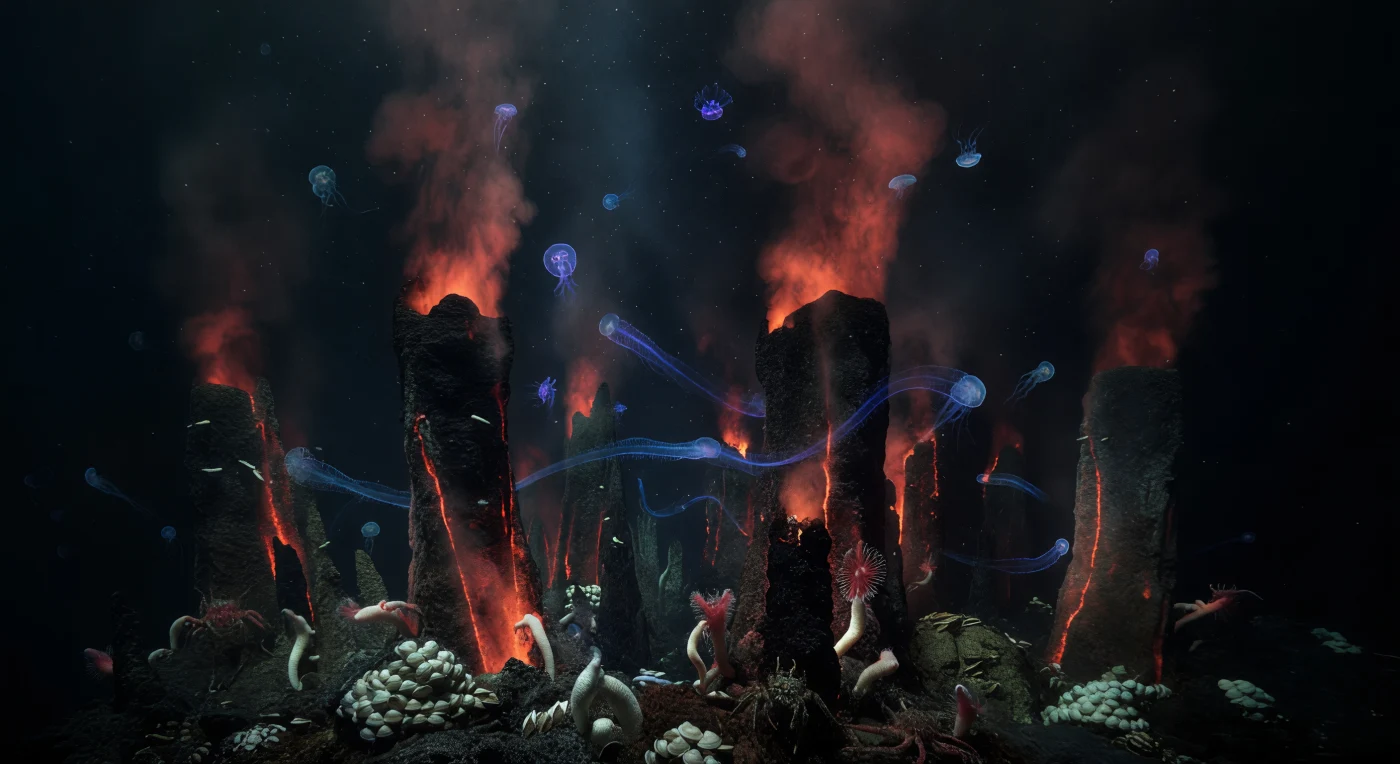
From fissures in fresh basalt along a mid-ocean ridge, superheated water charged with dissolved minerals erupts into the surrounding cold abyss, precipitating towering sulfide spires and black smoker chimneys that rise through pressures exceeding two hundred atmospheres — a world where tectonics and chemistry sustain life in complete sunless isolation. The only illumination here is Earth's own: faint chemiluminescent glows tracing the edges of newly formed mineral crusts, and the living phosphorescence of gelatinous animals drifting above the spire field in the open bathypelagic water column. Comb jellies and medusae pulse through the mineral haze, their bodies nearly transparent, made visible only by the blue, violet, and cyan bioluminescence their own tissues generate — trailing luminescent calligraphy through water that has not known sunlight for millennia. At the base of the chimneys, dense chemosynthetic communities — mats of sulfur-oxidizing bacteria, clusters of Calyptogena clams, pale Riftia tubeworms drawing sulfide directly from the vent flux through their trophosome organs, and slow-moving yeti crabs harvesting microbial films from mineral terraces — testify to an entire food web built on hydrogen sulfide rather than solar energy. Fine marine snow and mineral particles settle in absolute silence through this pressurized darkness, and the spires disappear upward into blackness, primordial and continuous, existing as they have long before any eye was present to perceive them.

At depths where sunlight has long since been extinguished and pressures exceed two hundred atmospheres, a black smoker chimney rises from the seafloor like a monument to Earth's interior heat, its fractured sulfide walls radiating superheated fluid at over 350 degrees Celsius into water only a few degrees above freezing. The chimney face glows with a dim copper-orange chemiluminescence where mineral-rich fluid meets the cold ocean column, fresh anhydrite crusts catching a faint ember warmth along their edges while iron sulfide deposits blister and oxidize into rust-orange stains across the matte black rock. Dense colonies of Pompeii worms — Alvinella pompejana, among the most heat-tolerant animals known to science — pack every crevice and mineral tube along the chimney wall, their pale segmented bodies anchored deep within their papery tubes while feathery crimson gill plumes ripple in the turbulent vent effluent, hosting a coat of thermophilic bacteria that may itself insulate them from the extreme thermal gradient. Around them, sparse bioluminescent pinpricks of cyan and blue-green pulse from other vent-associated organisms, and a slow dark plume of mineral particulates drifts upward into absolute blackness, dispersing into a water column where chemosynthesis — not photosynthesis — underpins an entire food web independent of the sun. This is a world powered entirely from within the planet, alive and intricate, existing in complete, pressurized silence long before any eyes could witness it.

Along a narrow rift cutting through freshly extruded basalt, superheated fluids charged with hydrogen sulfide and dissolved minerals surge upward through black smoker chimneys, precipitating towering mineral spires that exhale dense, particle-laden plumes into water held at crushing pressures between one hundred and four hundred atmospheres. The fracture itself retains a muted ember-glow where new crust has cracked open, and chemosynthetic microbial mats coat the surrounding rock in pale ghostly films, forming the base of an ecosystem that owes nothing to sunlight — here, energy flows from the planet's interior outward through chemolithotrophic bacteria and the Riftia tube worms and yeti crabs that graze upon them. Through this volcanic architecture drifts a gulper eel, Eurypharynx pelecanoides, its distensible jaw — evolved to engulf prey far larger than itself — held open as it moves through the absolute dark, its threadlike tail dissolving into the surrounding abyss as though the animal is being gradually unmade by the pressure and blackness. Faint bluish-green bioluminescent pulses from drifting zooplankton punctuate the dark above the ridge, while marine snow — the slow, continuous fall of organic detritus — descends through water that has never known a photon of solar origin. This is a world assembled entirely from heat, chemistry, and geological chance, silent and self-sufficient, indifferent to any observation.

At roughly one to four kilometres below the surface, where pressure exceeds one hundred atmospheres and the last trace of solar light dissolved long ago, a viperfish hangs in the water column as a pure silhouette — its needle fangs and elongate body lit only by the diffuse orange-red chemiluminescent glow rising from the black-smoker field far beneath it. Down on the basalt seafloor, hydrothermal chimneys exhaust superheated fluid at temperatures approaching 350 °C, precipitating towering sulfide structures and wreathing the surrounding rock in mineral-laden plumes; the thermal gradient sustains dense chemosynthetic ecosystems — bacterial mats, Riftia pachyptila tubeworm colonies with their crimson plumes, carpets of Calyptogena clams, and pale yeti crabs pressing against warm sulfide ledges — all of them drawing energy from hydrogen sulfide rather than sunlight. Scattered through the open water above, cyan and blue-green bioluminescent pinpricks mark drifting organisms too small to name individually, their faint emissions the only illumination beyond the vent glow itself. Marine snow and fine mineral particles drift slowly downward, carrying the chemistry of the mid-water into this benthic world, while the viperfish — Chauliodus sloani or a close relative, a pursuit predator of the midnight zone — remains perfectly still for an instant, suspended between the black water above and the primordial heat below, belonging entirely to a world that has existed on these terms for millions of years.

At mid-ocean ridge depths where tectonic plates pull apart and fresh basalt wells up from the mantle, mineral-laden hydrothermal fluid surges through a corroded sulfide archway — a monument built entirely by chemistry, precipitated over decades as superheated water carrying hydrogen sulfide, iron, copper, and zinc met the near-freezing abyssal water column and crystallized into these baroque, laminated structures. Beyond the arch, black smoker chimneys exhale plumes at roughly 350°C, the violent thermal gradient creating shimmering heat distortion as dissolved minerals instantaneously precipitate into dark, billowing clouds that rise and disperse into seawater held near two degrees Celsius under pressures exceeding 300 atmospheres. Dense colonies of pale limpets and Calyptogena clams cling to the shadowed sulfide base, their tissues nourished not by sunlight but by endosymbiotic bacteria that oxidize hydrogen sulfide through chemosynthesis — the same biochemical logic that sustains Riftia pachyptila tube worms and Yeti crabs clustered closer to the active vent effluent, each organism calibrated to exploit a narrow thermal and chemical gradient. A faint chemiluminescent and thermal glow pulses within the plume itself, and sparse bioluminescent points drift through the surrounding darkness like embers scattered in cold water, the only illumination in a world that has never known the sun and does not require it.

Along a fractured terrace of fresh black basalt on the mid-ocean ridge, the boundary between life strategies is written in shell and mineral: broad beds of white clams occupy the cooler sediment margins where chemosynthetic bacteria thrive within their tissues, while dense mussel aggregations pack tightly along the thermal gradient closer to active fissures, their dark satin shells overlapping in layered ranks. Above the seafloor, refractive shimmer betrays the mixing of superheated, mineral-laden fluid with near-freezing abyssal water, a boundary where hydrogen sulfide and methane fuel the entire food web through chemosynthesis rather than any trace of sunlight — which has not reached this depth, somewhere between one and four kilometers down, for as long as this ridge has been splitting apart. Tall black smoker chimneys exhale billowing plumes of three-hundred-and-fifty-degree fluid laden with iron, zinc, and copper sulfides, their charcoal columns dissolving into the immense cold darkness above, while a faint orange-red chemiluminescent glow pulses within the hottest effluent and a ghostly sheen drifts across pale microbial mats coating the surrounding crust. A yeti crab, its feathery setae dense with chemosynthetic bacteria, sits motionless among the mussels, farming its own food at a pressure exceeding two hundred atmospheres, while tiny cyan pinpricks of bioluminescence trace the path of small scavengers moving along the clam-to-mussel transition. Beyond the vent field, the water collapses into pure black — a primordial silence that has never required a witness.

At depths where pressure exceeds two hundred atmospheres and the last trace of solar radiation vanished kilometers above, stepped terraces of fresh basalt host one of the ocean's most improbable ecosystems. Diffuse venting breathes warm, mineral-laden fluid upward through cracks and fissures, sustaining thick white microbial mats that drape each volcanic ledge like sodden silk, their fringed margins trembling in a pearly haze of precipitating silica and anhydrite. Chemolithoautotrophic bacteria at the base of this food web oxidize hydrogen sulfide to fix carbon in total darkness, feeding dense aggregations of white Calyptogena clams wedged between mat-covered ledges, pale Riftia pachyptila tube worms rising stiffly from fissures, and Yeti crabs — Kiwaidae — crowding the warmer margins of the terrace, their feathery chelipeds harvesting the microbial film. Farther across the field, low chimneys exhale shimmering plumes whose turbulent edges glow with the faint orange-red chemiluminescence generated by superheated fluid meeting cold seawater, while newly fractured basalt carries a thermal ember of its own, and scattered cyan-blue-green points of bioluminescence pulse from pressure-adapted organisms drifting in the near-bottom water. This entire world — geological, chemical, and biological — runs on heat from the planet's interior alone, indifferent to the surface, sustaining itself in permanent night.

At depths where crushing pressure renders the water column an absolute void of sunlight, a black-smoker chimney of iron and copper sulfides rises from freshly erupted basalt along a mid-ocean ridge, its active orifice venting superheated fluid at roughly 350 °C directly into near-freezing abyssal seawater — a thermal and chemical collision that precipitates the dense black plume billowing upward in folded, stratified turbulence. Around the orifice, a dense living halo of *Rimicaris* vent shrimp crowds the mineralized rock in concentric layers, their translucent exoskeletons catching the faint orange-red chemiluminescent glow emanating from within the plume itself, where oxidizing sulfide compounds release trace wavelengths of light detectable only at this primordial scale. These shrimp are functionally blind in the conventional sense yet possess dorsal photoreceptors adapted to perceive precisely this geothermal radiance, navigating the thermal gradient between lethal scalding and the surrounding 2 °C abyss with evolutionary precision honed across millions of years. The vent ecosystem operates entirely beyond photosynthesis, sustained instead by chemolithoautotrophic bacteria that oxidize hydrogen sulfide into biomass, forming the base of a food web that requires nothing from the sun — a sovereign biosphere thriving at pressures exceeding 300 atmospheres, in permanent darkness, as it has since long before any witness existed to observe it.

At roughly 2,500 metres below the surface, where pressure exceeds 250 atmospheres and no solar photon has ever penetrated, a black smoker chimney exhales superheated fluid at 350 °C into near-freezing ambient water, precipitating a continuous fall of metallic sulfide grains — iron pyrite, chalcopyrite, anhydrite — that drift like volcanic ash over a narrow ledge of fresh basalt. Squat lobsters of the genus *Munida* or *Pleuroncodes* cluster along the mineral-crusted rock, their pale carapaces dusted with fresh precipitate, feeding opportunistically on microbial mats and organic particles carried by the plume's convective upwelling, while broad-bodied scale worms press flat against the sulfide surface, their iridescent paleae catching the only available light — a faint ember-orange chemiluminescence generated within the hottest vent effluent by thermochemical reactions, and dim cyan-green bioluminescent pulses from microbial communities colonising the chimney walls. Beyond the plume's thermal halo, the water reverts to its true nature: black, cold, utterly silent, threaded only by the slow vertical drift of marine snow descending from a sunlit world thousands of metres above. Here the entire food web inverts the logic of the biosphere, drawing energy not from the sun but from the oxidation of hydrogen sulfide by chemolithoautotrophic bacteria, sustaining an ecosystem of ghostly white clams, vestimentiferan tube worms, and crustaceans that have never needed light to thrive — a sealed volcanic economy operating in complete indifference to the illuminated ocean above.

Where the mid-ocean ridge tears itself apart along a great rift valley, superheated fluid surges upward through towering sulfide chimneys at temperatures exceeding 350 °C, billowing chemically rich plumes that bend and warp the surrounding water in transparent, heat-mirage layers — reality itself appearing to ripple across the basalt floor. At pressures approaching 400 atmospheres, sunlight is a distant irrelevance; here, energy flows not from the sun but from the planet's own interior, sustaining entire ecosystems through chemosynthesis, where microbial communities fix carbon from hydrogen sulfide and support cascading food webs entirely independent of the photic world above. Dense colonies of Riftia pachyptila extend vivid crimson plumes from their pale tubes, filtering dissolved compounds from vent effluent, while white clams settle into calmer sediment pockets and pale yeti crabs cling to chimney ledges crusted in iron sulfides and anhydrite, their hairy chelipeds harvesting chemosynthetic bacteria. Across the broader valley floor, freshly fractured pillow lavas retain a faint ember warmth in their narrow fissures, their glassy surfaces reflecting scattered cyan-green bioluminescent sparks from drifting organisms whose light signals have no witness here. Marine snow and fine mineral particulates drift silently through the abyssal water column, and the background dissolves into true oceanic blackness — a primordial darkness that has persisted, indifferent and complete, long before any eye existed to perceive it.

Where tectonic plates pull apart along a mid-ocean ridge, seawater percolates kilometers into fractured basalt, superheats against magma, and surges back as mineral-laden fluid at temperatures exceeding 350 °C — and from this violent chemistry, an entire ecosystem assembles itself in permanent darkness. Dense groves of *Riftia pachyptila* rise in pale ivory columns, some exceeding two meters, their vivid scarlet plumes — rich with hemoglobin adapted to carry both oxygen and hydrogen sulfide simultaneously — hovering in the milky shimmer of diffuse vent flow as it washes upward through cracked sulfide crust and jagged basalt. White bacterial mats blanket the warmest rock surfaces, fixing energy through chemosynthesis rather than sunlight, forming the photosynthesis-independent foundation of a food web that sustains clusters of *Calyptogena* clams wedged into seafloor pockets and pale yeti crabs (*Kiwa* sp.) braced at the scalding periphery of the colony, their long setae combing chemosynthetic bacteria directly from the superheated gradient. Black smoker chimneys exhale dense columns of precipitating minerals — anhydrite, pyrite, chalcopyrite — against water held at roughly 2 °C and 300 atmospheres of pressure, while the newest basalt seams carry the faintest thermoluminescent warmth along their fracture lines. Beyond the vent garden's haze of heat distortion and suspended mineral particles, the surrounding seafloor drops away into a darkness so absolute and so pressurized that it constitutes its own geological silence — a world that has sustained itself, without sunlight, for millions of years.

Where the mid-ocean ridge fractures open along fresh basalt, superheated fluid — laden with dissolved metals, sulfides, and silicates — erupts upward through towering chimney structures at temperatures exceeding 400 °C, billowing into ambient seawater barely above freezing and precipitating an incessant mineral snow: sulfide crystals, anhydrite flakes, and bacterial aggregates that drift slowly downward through absolute darkness, caught only by the faint chemiluminescent ember-glow radiating from within the plumes themselves. At pressures between 200 and 400 atmospheres, no sunlight has penetrated this water for millennia, yet the ecosystem here is extravagantly alive — dense stands of *Riftia pachyptila* tube worms extend crimson gill plumes into the shimmering, heat-distorted water column, their tissues nourished not by photosynthesis but by endosymbiotic chemoautotrophic bacteria that oxidize hydrogen sulfide as an energy source. Ghost-white *Calyptogena* clams crowd the warm cracks in the basalt, and pale yeti crabs — their chelipeds thickly colonized by filamentous bacteria — press against sulfide ledges glazed with microbial mat, harvesting the chemical gradient between the scalding effluent and the surrounding abyss. Scattered cold points of cyan and blue-green bioluminescence pulse among the fauna and the rising particulate, the only light that belongs entirely to life itself — a primordial economy of chemistry and darkness, cycling sulfur and carbon through a community that has no need of the sun, and no knowledge of the surface world far above.
