Scientific confidence: Very High

At roughly 2,500 metres below the surface, where pressure exceeds 250 atmospheres and the last photon of sunlight faded thousands of metres above, a freshly sunken whale transforms a barren silt plain into one of the ocean's most violent feasts. The carcass sprawls immense on the seafloor, its torn flanks exposing ivory slabs of blubber and deep crimson muscle, and the surrounding water erupts in cold cyan-blue bioluminescence wherever disturbed scavengers collide with flesh and sediment — each flash briefly tracing the waxy folds of skin, the gouged sediment, the writhing knots of hagfishes burrowing deep into the wounds. Sleeper sharks, heavy-bodied and scarred, wheel through the darkness in slow arcs, their slate-gray mass emerging and dissolving at the edges of the living light as they tear strips from the exposed flank, their biology tuned to extreme cold and near-zero oxygen metabolism. Clouds of fine silt drift upward from the churned seafloor, merging with the perpetual marine snow descending from the water column above, as countless smaller scavengers boil across the carcass edge, each disturbance triggering another pulse of bioluminescent light that maps the scene and then retreats into absolute black. This is the first stage of a succession that will bind this patch of seafloor for decades — the mobile-scavenger feast that precedes enrichment, bacterial mats, and eventually a chemosynthetic reef built on the mineral architecture of the bones themselves.
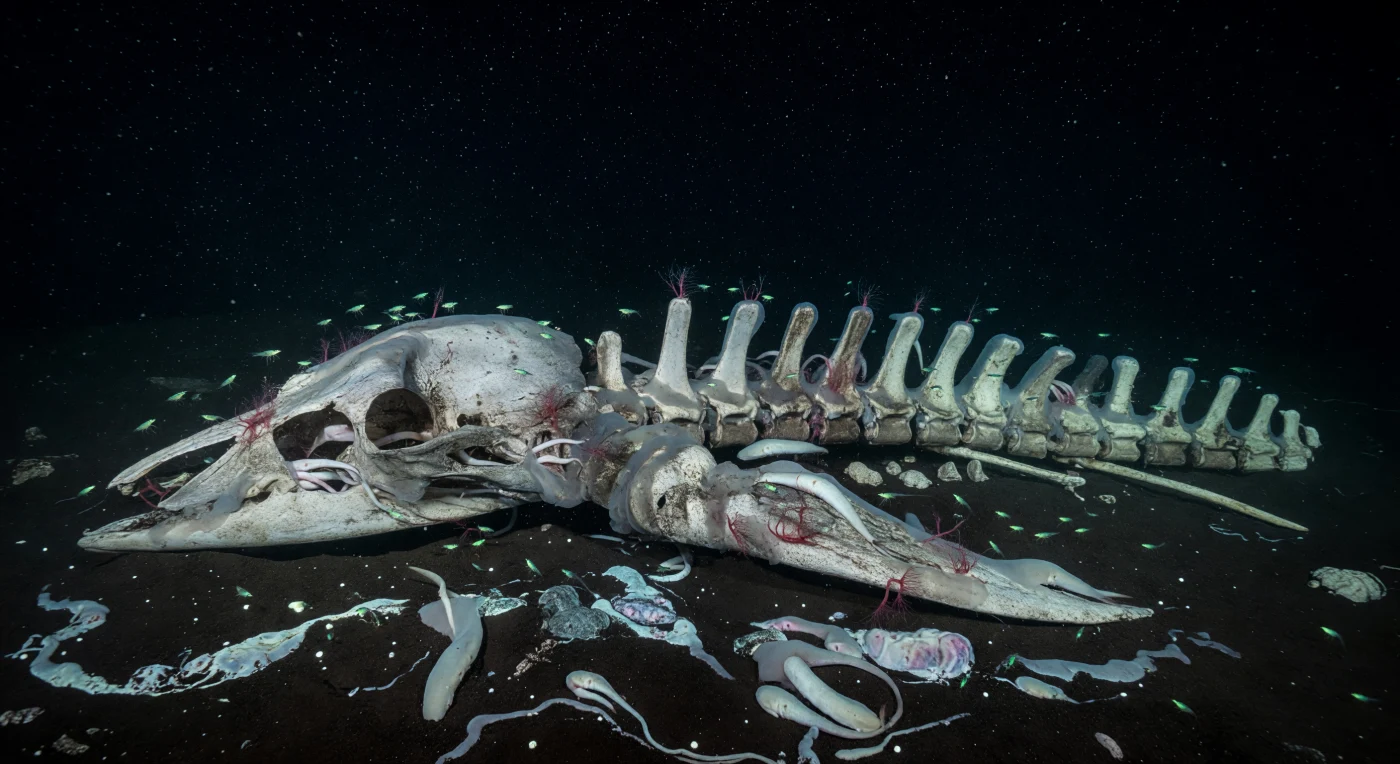
At depths where pressure exceeds two hundred atmospheres and no photon of sunlight has ever reached, a whale skull and its long chain of vertebrae rest half-buried in anoxic sediment, becoming an entire world unto themselves. The carcass has passed through the violent scavenger stage and now exists in its chemosynthetic afterlife — the bones themselves are the substrate, their lipid-saturated interiors exhaling hydrogen sulfide through slow anaerobic bacterial decomposition, sustaining a reducing microhabitat in the midst of an otherwise impoverished abyssal plain. Osedax worms, the so-called bone-eating snot-flowers, anchor their root-like tissues directly into the porous matrix of each vertebra, extracting collagen with the help of endosymbiotic bacteria, their crimson gill plumes rising in delicate fans from every available surface while hagfish coil through the orbital cavities of the skull in slow, muscular loops. The bacterial carpets draped across the ivory and ash-gray bone are not passive decay but active chemosynthetic communities, converting sulfide to biomass just as their cousins do at cold seeps thousands of kilometers away, and the emerald flickers tracing each ridge belong to small shrimp grazing these microbial films — brief cold sparks of bioluminescence in water that carries marine snow silently downward through absolute darkness. This is the long chemical afterlife of a giant, a decades-long cascade of ecological succession playing out in perfect silence, with no witness, in water so cold and still it preserves every filament of bacterial velvet exactly where chemistry placed it.

At depths where pressure exceeds three hundred atmospheres and sunlight has been absent for centuries, a whale's skeleton transforms the otherwise barren abyssal mud into a thriving, self-sustaining metropolis of life. The pale arching ribs, stripped by months of scavenger activity, rise from the dark sediment like the nave of a sunken cathedral, their surfaces coated in sulfurous bacterial films fed by lipids still seeping from within the bone matrix — a chemosynthetic process that mirrors, in miniature, the biology of cold seeps and hydrothermal vents. Immense swarms of lysianassid amphipods pulse across the bones in cascading green-blue waves of bioluminescence, their brief cold flashes illuminating slick remnants of collagen, the crimson plumes of *Osedax* worms boring silently into the smaller bones to digest their fats, and hagfish threading through the darkened cavities of the skeleton in slow, boneless loops. Beyond the colonnade, barely resolved in the absolute blackness of the water column, the silhouette of a Greenland sleeper shark drifts at the periphery — metabolically glacial, unhurried, drawn by chemoreception across kilometers of featureless mud to this improbable abundance. Marine snow descends in all directions through the stillness, each particle a fragment of the surface world finally arriving here, joining an ecosystem that will persist and transform for decades without ever requiring the presence of light.

At roughly 2,500 meters beneath the surface, where sunlight has been absent for centuries and the water presses down at pressures exceeding 250 atmospheres, a cathedral of bleached bone rises from the soft abyssal sediment — all that remains of a great whale that sank months or years ago. The skeleton is thick with life: pale bacterial sulfide mats spread across the vertebrae like frost, Osedax worms bore silently into the lipid-rich bone matrix with root-like extensions, and hagfish coil through marrow cavities in slow, deliberate loops, while Greenland sleeper sharks circle the outer ribs with the measured patience of animals shaped by millions of years of cold and pressure. Above the skeleton, comb jellies and hydromedusae drift in loose spirals through the marine snow — fragments of organic matter sinking from the world far above — their iridescent ctene rows and pulsing bells casting cobalt and violet halos that briefly illuminate the arc of each rib before fading back into absolute darkness. The only light in this scene is biological: living animals advertising, hunting, and existing in a darkness so complete that the bones themselves seem to glow faintly only where microbial films catch the ambient bioluminescence. This carcass functions as an ecological island, a chemosynthetic oasis that concentrates biomass and biodiversity across a succession spanning decades, connecting the ocean's great productive surface to its most lightless, pressure-crushed floor through the slow chemistry of decay.
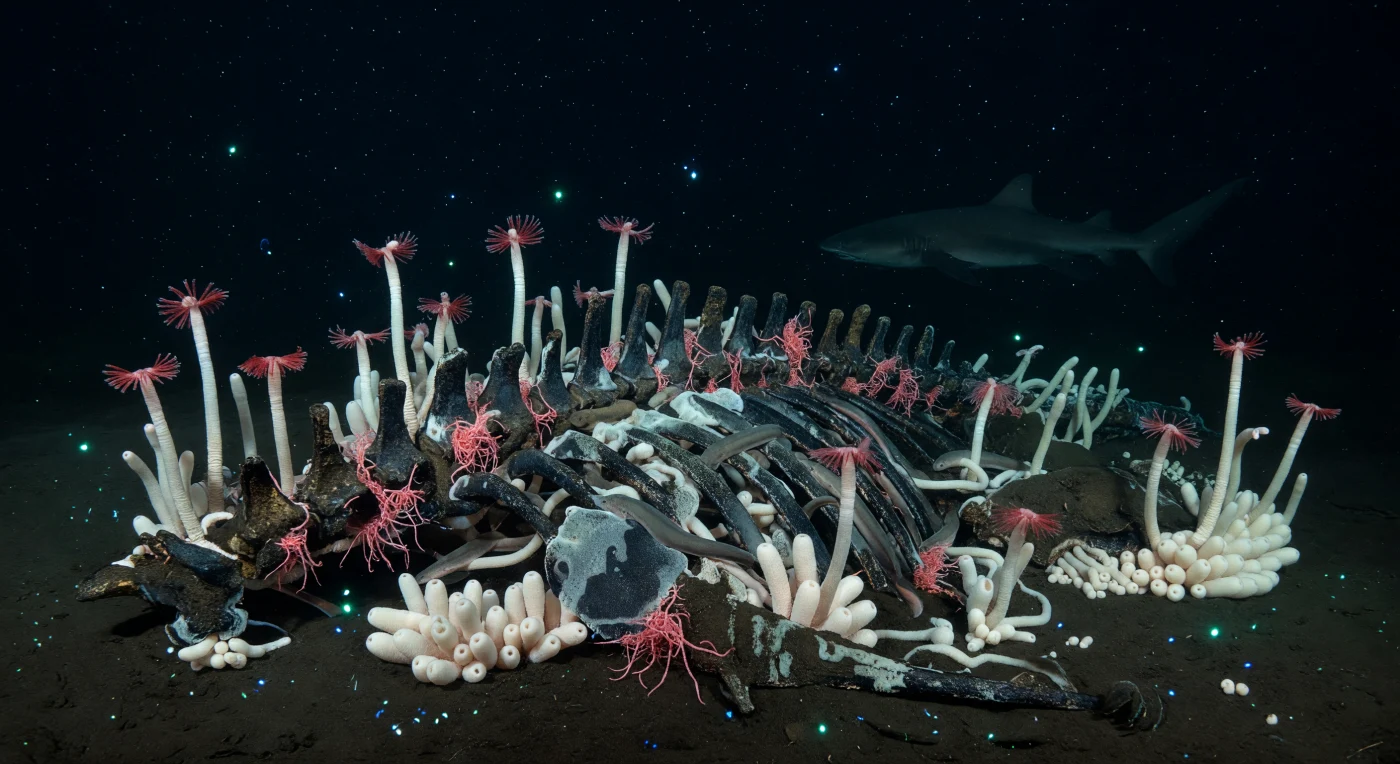
Where a great whale once drifted down through thousands of meters of darkness, its skeleton now anchors an entire world on the abyssal plain. At pressures exceeding 100 to 400 atmospheres and temperatures hovering near 2°C, the bones have become a chemical factory: anaerobic bacteria decompose lipid-saturated tissue within the vertebrae, generating hydrogen sulfide that diffuses upward through blackened sediment, feeding dense mats of sulfur-oxidizing microbes and sustaining a chemosynthetic garden that owes nothing to sunlight. From this sulfide-dark mud, ivory-sheathed tube worms rise in close-packed colonies, their crimson gill plumes — rich in hemoglobin specialized to bind both oxygen and hydrogen sulfide simultaneously — suspended motionless in water so cold and still it seems almost solid. Threadlike Osedax worms, the bone-devouring specialists discovered only in the early 2000s, bloom from exposed vertebral surfaces like pale rootlets, their digestive symbiont bacteria slowly dissolving the remaining skeletal matrix over years or decades. Intermittent pulses of cyan bioluminescence from drifting plankton wash briefly across wet bone and velvety bacterial film, then vanish, leaving this silent reducing ecosystem exactly as it has existed since the carcass settled — complete, indifferent, and entirely its own.

At roughly 2,500 metres below the surface, where pressure exceeds 250 atmospheres and no photon of sunlight has ever penetrated, a sperm whale skeleton sprawls across cold, silty sediment like the ruins of an ancient cathedral — vertebrae half-swallowed by the mud, the great rib cage arching upward in silent geometry. The fall has long since passed through its mobile-scavenger stage and entered the sulfophilic phase: pale, waxy mats of chemoautotrophic bacteria spread across bone surfaces, drawing energy not from light but from hydrogen sulfide generated as lipid-rich bone marrow decomposes beneath them, while Osedax worms — their feathery plumes flushed cream and red — bore directly into the porous vertebrae, hosting endosymbiotic bacteria that digest collagen and fat locked within. Through the calcified ribs, a scatter of sapphire and cyan flashes betrays startled amphipods and isopods streaming away from disturbance, their bioluminescent escape signals illuminating wet bone texture and drifting marine snow for fractions of a second before darkness reclaims everything. Crossing the midwater above this chemosynthetic island, a viperfish — Chauliodus sloani — cuts a near-perfect silhouette, its needle profile and recurved fangs evolved for a world of ambush and darkness, its passage utterly indifferent to the slow sulfurous transformation unfolding on the seafloor below. Here, far beneath the reach of seasons or storms, a single carcass sustains an entire archipelago of life across decades, chemosynthesis standing in for sunlight, bone replacing basalt.

At 2,500 metres beneath the surface, where pressure exceeds 250 atmospheres and no photon of sunlight has ever reached, a whale skeleton lies half-swallowed by the soft black silt of the abyssal plain, its great ribs arching upward like the nave of a drowned cathedral. The bones are deep in their third and final successional stage — the sulfophilic phase — carpeted in velvety chemosynthetic bacterial mats that harvest hydrogen sulfide seeping from lipid-rich marrow, hosting dense colonies of Osedax bone-worms whose feathery red and white plumes probe every surface with root-like rhizoids that dissolve the very matrix of the skeleton. A gulper eel — Eurypharynx pelecanoides — sweeps through the water column above in a slow arc, its body nearly dissolved into the darkness, only the vast unfurled membrane of its jaw catching form as a drifting cloud of ostracod bioluminescence detonates across the frame: hundreds of Conchoecia releasing simultaneous cyan sparks in a defensive cascade, briefly painting the silt ripples, the Osedax plumes, and the eel's translucent sail in cold turquoise fire. Hagfishes thread silently through the orbital cavities of the skull, and at the far margin of the fall the blunt silhouette of a sleeper shark idles with metabolic patience, its slow-oxidising biochemistry perfectly suited to a world where meals are separated by weeks and darkness is total, permanent, and immense.

At roughly 2,500 metres below the surface, where pressure exceeds 250 atmospheres and no trace of solar light has ever penetrated, a detached whale vertebra rests half-buried in fine abyssal sediment — a remnant of what was once a colossal carcass, now reduced to bone and chemistry. The skeleton has become an island of energy on an otherwise barren plain, its porous mineral surfaces colonised by a pale bacterial film stained faintly with sulfide, while nearby sediment hosts dense bacterial mats feeding on the organic compounds still seeping from decomposing lipid-rich bone tissue — the sulfophilic stage of whale-fall succession described by Smith and Baco. A female ceratioid anglerfish hangs motionless in the black water column beside the vertebra, her swollen body perfectly neutrally buoyant, her single bioluminescent esca emitting a cold cyan-green glow produced by symbiotic bacteria housed within the lure — the only illumination in this scene, catching needle teeth and drifting particles of marine snow that fall at less than a metre per day from the world above. At the edge of visibility, crimson Osedax plumes extend from another bone fragment, these bone-eating worms boring into the matrix with root-like tissue that digests collagen and lipids through symbiotic bacteria, binding the whale-fall community into a chemosynthetic web that owes nothing to sunlight. This is a world complete and ancient in itself, operating in permanent darkness, immense stillness, and cold near-freezing water, indifferent to any gaze.

At roughly 2,500 metres below the surface, where pressure exceeds 250 atmospheres and the last trace of solar light has long since ceased to exist, a cluster of ancient whale vertebrae forms the only hard substrate for kilometres across an otherwise featureless abyssal silt plain. These bones — porous, mineral-stained, and slowly dissolving across decades — represent the terminal stage of the Smith & Baco succession model: the sulfophilic phase, in which bacterial mats of filamentous chemolithotrophs spread across organically enriched sediment, oxidising hydrogen sulfide released by anaerobic microbial degradation deep within the lipid-saturated bone matrix, sustaining a chemosynthetic oasis entirely independent of sunlight. Ivory-toned anemones anchor their pedal discs into crevices of the vertebral arches, while pale ophiuroids — brittle stars — drape their long articulated arms across the bone rims, sweeping suspended particles from water so cold it hovers near 2°C and carries dissolved oxygen only in modest concentrations. From the water column above, a siphonophore drifts past in a slow, colonial procession, its coordinated bioluminescent pulses emitting brief cyan sweeps that trace the contours of ancient bone and translucent tentacle alike before the darkness reclaims everything. This reef of the dead sustains the living without sunlight, without warmth, and without any witness but the deep ocean itself.

At roughly 2,500 metres beneath the surface, the seafloor here is twin kingdoms sharing a single darkness: the slow archaeology of bone and the violent chemistry of the Earth's interior, held together by cold, pressure, and the complete absence of sunlight. A whale skull rests among shattered basalt talus, its calcium phosphate surfaces softened by velvet bacterial mats and threaded through by Osedax worms — bone-eating annelids whose root-like tissues secrete acid to dissolve the lipid-rich matrix within, extracting nutrition from skeletal material that may sustain a chemosynthetic community for decades under the Smith & Baco succession model. Pale hagfishes work the cavities with slow muscular coiling, and sleeper sharks — *Somniosus* species adapted to the crushing pressure of the midnight zone, where water column above compresses to over 250 atmospheres — describe wide, unhurried arcs around the carcass, their neutral buoyancy a biological solution to a world that offers no thermocline to soar upon. Beyond the bones, black smoker chimneys rise from fractured pillow basalt along an active spreading ridge, their anhydrite and sulfide mineral spires built incrementally by superheated fluid — exceeding 350°C at the vent mouth — that carries reduced iron, copper, and sulfur compounds into the near-freezing ambient water, generating orange-red chemiluminescent oxidation halos at the plume boundaries, while cyan and blue bioluminescent pulses from microbial films and small deep-sea fauna softly trace the skull's eroded sutures and the shark's skin in a world that illuminates only itself.
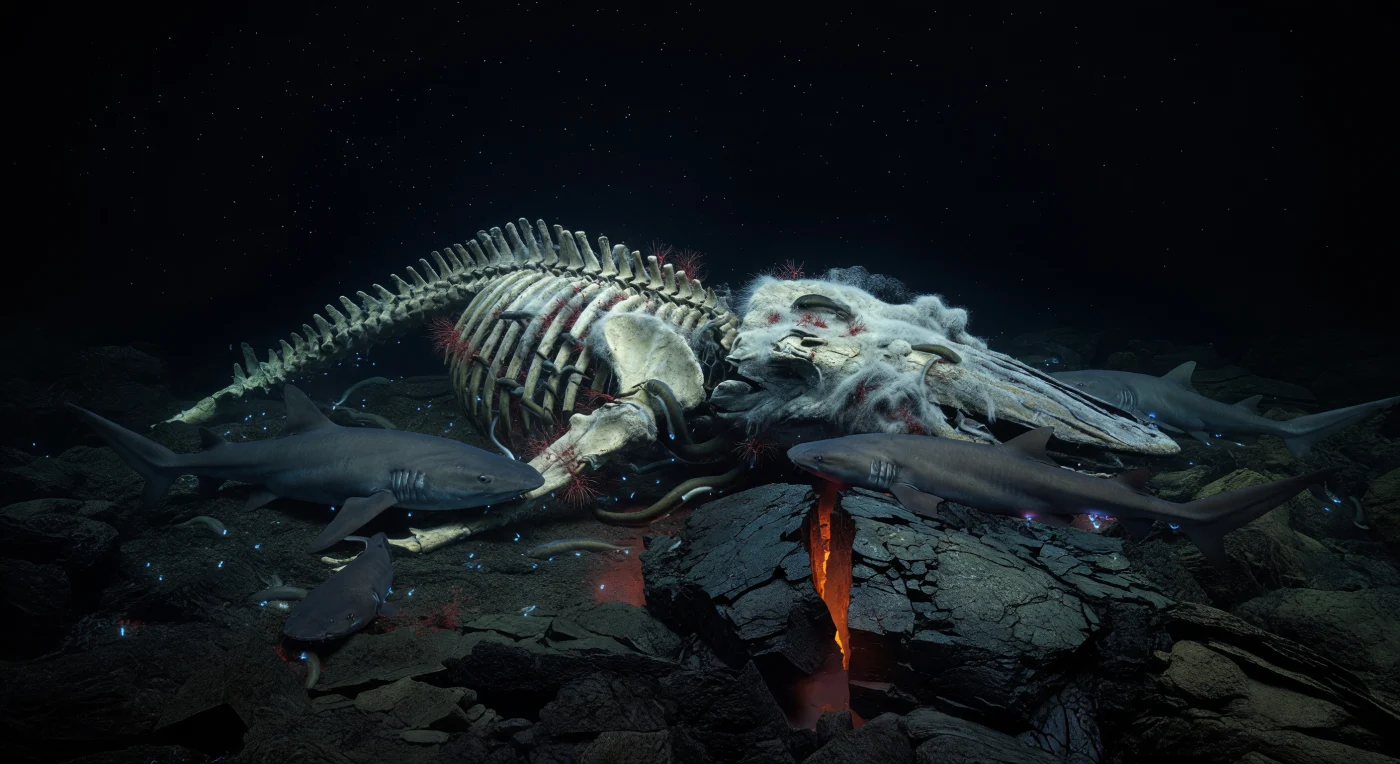
At depths where crushing pressure silences all but the chemistry of decay, a sperm whale's skeleton lies across a newly fractured basalt ridge, one of the few places on Earth where tectonic heat and biological richness converge without design. The fresh lava fracture still breathes a dull orange glow through its glazed black crust, casting mineral-dusted marine snow into faint relief as particles drift past razor-edged rock cooled seconds or centuries ago — time rendered visible in texture alone. Every exposed bone surface is colonized: vertebrae disappear beneath snow-white bacterial mats fed by sulfide rising from lipid-soaked sediment, Osedax worms bore silently into cortical bone while their feathery red plumes pulse with each chemical exchange, and hagfish twist through the orbital cavities and between collapsed ribs in slow, purposeful knots. Sleeper sharks — broad, unhurried, ancient in their patience — circle the periphery, occasionally pressing their blunt snouts into softened cartilage, their fin edges briefly outlined by the cold cyan sparks of bioluminescent organisms disturbed in the water column above. This carcass is not a site of death but of extraordinary productivity, a chemosynthetic oasis producing more biological complexity per square meter than almost any surrounding seafloor, its sulfide chemistry mirroring the metabolic logic of hydrothermal vents while the fresh magmatic heat beneath the basalt adds a geological pulse that no whale fall beside cold abyssal sediment ever possesses.

Where no sunlight has ever reached, a constellation of fractured vertebrae and arching ribs lies half-consumed by the seafloor itself, the bones bleached to chalk and furred with thick bacterial mats that glow with the faint, diffuse luminescence of sulfide-oxidizing microbial colonies. At roughly 2,500 meters, hydrostatic pressure exceeds 250 atmospheres, water temperature hovers near two degrees Celsius, and the chemical conditions within and around the decaying skeleton have shifted from aerobic decay to anaerobic sulfate reduction — a quiet geochemical revolution that transforms bone marrow into a hydrogen-sulfide factory sustaining an entire island ecosystem. From every crack and marrow channel, colonies of *Osedax* boneworms have anchored their colorless, root-like rhizoids deep into the lipid-saturated matrix, sending up hundreds of translucent stalks that bloom into vivid crimson gill plumes wavering motionlessly in the still, pressurized water, while white bacterial floc drifts free between the ribs like a perpetual slow snowfall. Hagfishes coil in the dark spaces between bones, their pale bodies slick against the sediment, and at the outermost edge of visibility a sleeper shark hangs suspended in the black water column, barely distinguishable from the void — a reminder that even in this most absolute of silences, the ocean's metabolism never stops.
