
In einer Tiefe zwischen 1.500 und 3.000 Metern, wo Tageslicht seit Jahrhunderten nicht mehr existiert und der Druck das Hundert- bis Dreihundertfache der Oberfläche erreicht, hängt der weibliche Tiefseeanglerfisch nahezu regungslos im schwarzen Pelagial – ein Tier, das die Evolution zur lebenden Falle geformt hat. Allein die Esca, jene biolumineszente Angelrute am verlängerten Dorsalstrahl, leuchtet in einem zartblauen Grünton, erzeugt durch symbiontische Bakterien, die in dem organischen Gewebe des Köders dauerhaft Licht produzieren: eine der raffiniertesten Jagdstrategien der Tiefsee. Im Schein dieses lebenden Lichts werden für Bruchteile eines Moments transluzente, nach innen gebogene Zähne sichtbar, perfekt konstruiert, um Beute, die einmal das Maul berührt, nicht mehr zu entlassen, während das samtschwarze Gewebe des Körpers sofort wieder in die totale Dunkelheit versinkt. Vereinzelte Partikel marinen Schnees – absinkende organische Fragmente aus den Schichten weit darüber – treiben lautlos vorbei und bilden die einzige Bewegung in diesem pressurisierten, nahezu stillstehenden Wasser von kaum mehr als zwei bis drei Grad Celsius. Diese Welt existiert in vollständiger Stille und vollständiger Eigenständigkeit, gehorcht nur den Gesetzen der Chemie, des Drucks und des biologischen Lichts – unberührt, urzeitlich und gleichgültig gegenüber allem, was außerhalb ihrer Dunkelheit liegt.

Im offenen Mittelwasser zwischen 1.500 und 2.500 Metern Tiefe, wo der hydrostatische Druck rund 150 bis 250 Atmosphären beträgt und kein Sonnenstrahl jemals eindringt, entfaltet sich ein Jagdaugenblick von äußerster Präzision: Ein Viperfisch — *Chauliodus sloani* — durchschneidet in schräger Stoßbewegung einen lockeren Schwarm tiefseepelagischer Krebstiere, seinen schmalen, pistolenblauen Körper von sporadischen biolumineszenten Zyanblitzen gezeichnet, die sich in Wellen durch das aufgeschreckte Schwarmgefüge ausbreiten und wieder erlöschen. Die Beleuchtung ist ausschließlich biologischer Natur — intermittierende Lichtpulse biolumineszenter Planktonorganismen und die Eigenleuchte des Fisches selbst —, die seine nadelspitzen Unterkieferzähne und seine großen, reflektierenden Augen für Sekundenbruchteile aus der Schwärze herauslösen, bevor die absolute Finsternis alles zurückfordert. Mariner Schnee, feine organische Partikel aus dem biologischen Kreislauf der oberen Wassersäule, treibt gleichmäßig durch das eiskalte Wasser von etwa 3 °C, ein steter Materialfluss, der die Energieversorgung der gesamten bathypelagischen Lebensgemeinschaft speist. Der Viperfisch besitzt einen ausgeprägten Leuchtorgan-Streifen entlang der Ventralseite — ein Gegenbeleuchtungssystem zur Tarnung gegenüber aufwärts blickenden Räubern — sowie ein hochgradig anpassungsfähiges Maul- und Kiefergelenk, das es ihm erlaubt, Beute von nahezu eigenem Körpermaß zu verschlingen, eine evolutionäre Antwort auf die extreme Nahrungsunvorhersehbarkeit dieser Zone. In diesem grenzenlosen Dunkel, weit von jedem Lichtschimmer und jeder Küste entfernt, existiert eine Welt vollständiger Stille und ununterbrochener Urzeitlichkeit, die sich selbst genügt und seit Jahrmillionen ohne Zeugen pulsiert.

In einer Tiefe, in der kein Sonnenstrahl je ankommt und der hydrostatische Druck mehrere hundert Atmosphären beträgt, schwebt ein Schwarzer Drachenfisch (*Idiacanthus* sp.) reglos im offenen Pelagial – ein langgezogener, samtschwarzer Körper, der fast vollständig im Nichts aufgeht. Entlang seiner Flanken senden Reihen von Photorophoren ein gedämpftes Cyansignal aus, das weniger der Jagd als der innerartlichen Kommunikation dient, ein biochemisches Erkennungszeichen in absoluter Dunkelheit, das von keiner anderen Lichtquelle gestört wird. Das fadenförmige Kinnbarbel hängt unterhalb des Kiefers, bestückt mit lumineszierenden Organen, die kleine Beutetiere aus der lückenhaften Nahrungsmatrix des Bathypelagials anlocken – einem Lebensraum, in dem Beute selten und unberechenbar ist und jede Begegnung entscheidend sein kann. Winzige Partikel marinen Schnees driften lautlos durch das kalte, klare Wasser und werden nur dort sichtbar, wo das schwache lebende Leuchten des Fisches sie für einen Augenblick aus der Schwärze löst. Dieser Ozean existiert vollständig in sich selbst – ohne Zeuge, ohne Oberfläche, ohne Boden, ein druckvolles Schweigen, das seit Jahrmillionen andauert.

In einer Tiefe, wo das Sonnenlicht längst erloschen ist und der hydrostatische Druck auf mehrere hundert Atmosphären angewachsen ist, gleitet ein Koboldhai in vollkommener Stille über den Abbruch einer basaltischen Steilwand — sein abgeflachtes, verlängertes Rostrum, die blasse, fast durchscheinende Haut und das weiche aschgraue Muskelgewebe kaum vom umgebenden Schwarzwasser zu unterscheiden, nur angedeutet durch einen letzten, kaum messbaren Rest blauen Abwärtslichts, das aus Tausenden Metern Wassertiefe herabgefiltert wurde. Scattered biolumineszente Funken — Cyan- und Blaugrün-Punkte von Plankton und kleinen Organismen — treiben wie lebendige Partikel durch die Wassersäule, umrahmt von feinem marinen Schnee, der lautlos in dieser thermisch stabilen Welt von zwei bis vier Grad Celsius sedimentiert. Der Hai, mit seinen druckangepassten, schwach mineralisierten Geweben und seinem gelatinösen, mechanisch effizienten Körperbau, hängt reglos über Stufen aus gebrochenem Vulkangestein, dessen frische Basaltoberflächen und mineralische Krusten Zeugnis tektonischer Aktivität sind — ein Ökosystem, das in seiner heterogenen Dunkelheit eigenen Gesetzen folgt. Dieser Raum existiert jenseits jeder menschlichen Wahrnehmung, ein uraltes Raubtier in einem Reich, das sich selbst genug ist — still, kalt, verdichtet und unendlich gleichgültig gegenüber allem, was oberhalb seiner Schwelle geschieht.

In den lichtlosen Tiefen zwischen tausend und viertausend Metern hängt ein Magnapinna-Tintenfisch reglos in der Wassersäule, sein durchscheinender Mantel und die breiten, dreieckigen Flossen kaum fassbar im filigranen Leuchten fernen Planktons — ein lebendiges Kaltlicht aus Cyan und Blau, das keine Wärme kennt und keinen Schatten wirft. Die Arme dieses rätselhaften Kopffüßers, so lang wie kaum ein anderes Tier sie trägt, knicken in jenen charakteristischen, ellbogenartigen Winkeln ab und verlieren sich in der unermesslichen Schwärze darunter, als wären sie Fäden, die das Tier an einer unsichtbaren Tiefe verankern. Der hydrostatische Druck übersteigt hier zweihundert Atmosphären, das Wasser ist wenige Grad über dem Gefrierpunkt, und Meeresschnee — die rieselnden Überreste des Lebens von oben — treibt lautlos an transparenten Geweben vorbei, die sich über Jahrmillionen an eine Existenz ohne Licht, ohne Boden und ohne Zeugen angepasst haben. In dieser bathypelagischen Mitternachtszone gilt weder Schwerkraft noch Stille als Metapher: Sie sind physische Realitäten, in denen Organismen wie dieser Tintenfisch ihre Sinne, ihre Chemie und ihre Körper so verfeinert haben, dass Jagd, Überleben und Fortpflanzung im Verborgenen möglich werden. Der Ozean existiert hier vollständig in sich selbst — unbeirrt, unangetastet und von keinem Blick erwartet.
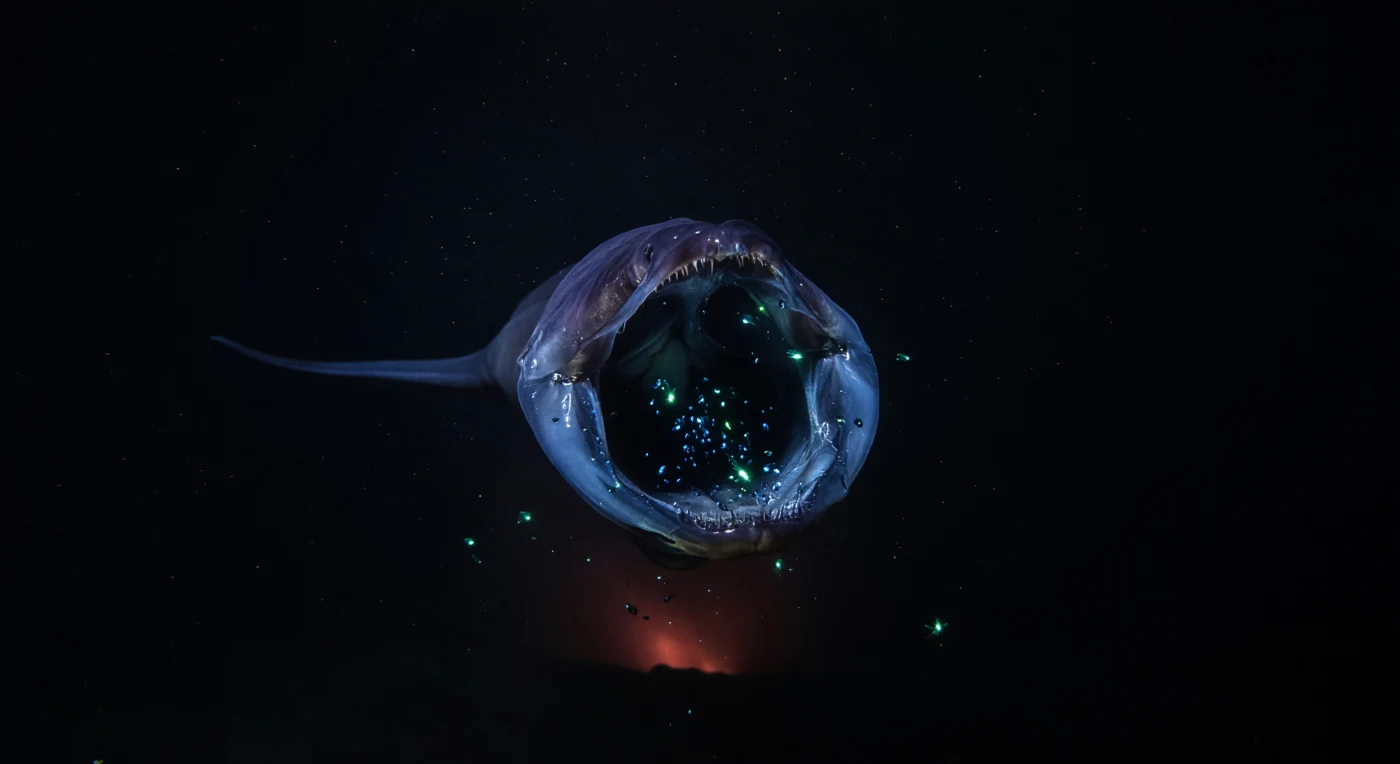
In einer Tiefe zwischen tausend und viertausend Metern, wo kein Sonnenstrahl je eindringt und der hydrostatische Druck mehrere hundert Atmosphären beträgt, vollzieht sich einer der radikalsten Fangreflexe der Tiefsee: Ein Schlundaal (*Eurypharynx pelecanoides*) reißt seinen membranartigen Kiefer zu einer transluzenten blauen Kuppel auf, die einen ganzen Schwarm winziger Beutetiere umschließt, deren panische Biolumineszenz das Innere des Rachens in kalten Cyan-Grüntönen aufleuchten lässt und nadelförmige Zähne sowie glitschige Gewebewände schemenhaft sichtbar macht. Das Tier selbst ist ein evolutionäres Paradoxon – ein Wirbeltier mit einem Schädel, dessen Gelenkmechanik alle anatomischen Proportionen aufgelöst hat, der Körper druckadaptiert, muskelschwach und dunkelviolett, der peitschartige Schwanz bereits im Nichts verschwunden. Weit unterhalb, kaum wahrnehmbar, färbt ein diffuser orangeroter Schimmer hydrothermaler Herkunft die tiefsten Bildschichten, zu schwach, um als Licht zu gelten, stark genug, um an die geochemische Aktivität des Meeresbodens zu erinnern. Mariner Schnee – organische Partikel, die langsam aus den oberen Wasserschichten herabsinken und die einzige Nahrungsbrücke zwischen Sonnenlicht und dieser absoluten Dunkelheit darstellen – driftet lautlos durch das Bild, während das Wasser selbst kalt, klar und gleichgültig bleibt gegenüber dem Ereignis, das es umhüllt.

Im Inneren des mittelozeanischen Rückens, Tausende Meter unter der lichtlosen Wasseroberfläche, sickert ein rötlich-oranges Glimmen durch Risse frisch erstarrter Basaltlava – ein geologisches Glühen, das aus dem Erdinnern selbst stammt und gegen das schwarze, nahezu eiskalte Wasser dringt, das hier unter einem Druck von mehreren Hundert Atmosphären lastend stillsteht. In dieser chemischen und thermischen Grenzlandschaft, wo hydrothermale Fahnen in chemolumineszenten Schleiern aufsteigen und das Wasser mit Mineralteilchen und Meeresschnee durchsetzt ist, bewegt sich ein Anglerfisch lautlos an den gebrochenen Lavaterrassen vorbei – sein blauweißes Leuchtorgan zittert wie ein falsches Sternchen vor einem Gebiss aus gebogenen, durchsichtigen Zähnen, während biolumineszierende Planktonorganismen seinen Körper in Cyan und Grün aufblitzen lassen. Tiefer im Dunkel zeichnet sich das Silhouett eines Viperfischs ab, sein metallisch-dunkles Schuppenkleid reflektiert das vereinzelte Biolumineszenzflackern, während die fadenförmigen Tentakelanhänge eines Magnapinna-Tintenfischs wie unmöglich dünne, blasse Bänder vertikal in die Wassersäule hinabfallen. Diese Geschöpfe sind Meister einer Existenz, die von extremer Drucktoleranz, biochemischer Anpassung und der evolutionären Perfektion der Stille lebt – eine Welt, die kein Tageslicht kennt und keine Zeugen braucht.

In einer Tiefe von mehr als zweitausend Metern steigen die Schlote aus frischem, dunklem Basalt empor und stoßen dichte, mineralreiche Fahnen aus, die sich wie Säulen aus verfestigter Dunkelheit in das nahezu vollständige Schwarz des umgebenden Wassers ergießen — ein Prozess, der seit Jahrmillionen ohne Zeugen abläuft. An den heißesten Austrittsöffnungen, wo überhitztes, schwefelbeladenes Fluid auf nahezu gefrierendes Tiefenwasser trifft und Temperaturgefälle von Hunderten von Grad über wenige Zentimeter entstehen, zittert ein rostroter bis orangefarbener Schimmer, der einzige Widerschein thermischer Energie in dieser absoluten Finsternis. Ein bathypelagischer Raubfisch gleitet mit präziser, druckadaptierter Muskulatur entlang des Fahnenrands, sein Umriss von vereinzelten blaugrünen Biolumineszenzpunkten aufgelöst — lebendige Lichter, die in einem Ozean ohne Sonne das einzige Gegengewicht zur Schwärze bilden. Tiefer im Bild deutet das winzige Lockorgan eines Tiefseeanglerfisches auf eine Strategie hin, bei der Licht nicht erhellt, sondern täuscht, während im Hintergrund die gestreckte Silhouette eines Viperfisches und die gespenstischen, endlos langen Arme eines Magnapinna-Tintenfisches lautlos im partikelreichen Wasser vergehen. Bei einem Druck von über zweihundert Atmosphären, in einer Welt, in der Nahrung knapp, Begegnungen selten und Sinne auf mechanische Präzision ausgelegt sind, existiert dieses Ökosystem vollständig in sich selbst — uralt, still und vollkommen gleichgültig gegenüber allem, was oberhalb seiner Reichweite liegt.

Im Grenzbereich eines diffusen Hydrothermalaustritts, wo mineralreiches, leicht erwärmtes Tiefenwasser auf nahezu gefrorenes Abgrundwasser trifft, verharrt ein Tiefseeanglerfisch in vollkommener Reglosigkeit – der Körper druckadaptiert, das Gewebe weich und nachgiebig bei annähernd 300 Atmosphären hydrostatischem Druck. Sein lumineszentes Lockorgan, ein konzentrierter blaugrüner Punkt aus bakteriell erzeugtem Biolicht, hängt reglos über schwefelblassen Mineralablagerungen und frisch gebrochenem Basalt, dessen glasige Kanten noch eine schwache thermische Eigenstrahlung abgeben. Um ihn herum pulsieren sporadische Biolumineszenzflicker der Ventfauna in Cyan und Grün, während ein gedämpfter orangegelber Chemolumineszenzschimmer knapp über dem Gestein aufsteigt – das gesamte schwache Licht ist biologischen und geochemischen Ursprungs, ohne jede andere Quelle. Mariner Schnee und feinste Mineralpartikel treiben träge durch das dichte, klare Wasser, unberührt von jeder Strömung außer dem kaum wahrnehmbaren Schleiern des Ventaustriebs, der die Grenzschicht zwischen warm und kalt als zitternde Brechungsverzerrung sichtbar macht. Hinter dem Räuber verschluckt die absolute Dunkelheit jeden Kontext, unterbrochen nur von vereinzelten fernen Biolumineszenzpunkten – Hinweise auf Leben jenseits des Ventrands, das seit Jahrmillionen ohne Zeugen existiert.

In einer Tiefe, wo der Druck bis zu vierhundert Atmosphären beträgt und kein Sonnenstrahl je eindringt, liegt ein Walgerippe auf dem feinen dunklen Sediment des Bathypelagials – ein gestrandeter Überfluss inmitten ewiger Knappheit, der die Nahrungsökologie des Mitternachtsmeeres für Jahrzehnte umstrukturiert. Cyan-grüne und blaugrüne Biolumineszenzblitze zucken durch die Wassersäule, wenn Aasfresser und Räuber in überlagerten Schwärmen über den Knochen kreisen: Viperfische hängen schräg im Schwarz, ihre Nadelzähne für den Bruchteil einer Sekunde von fremdem Lebendlicht gestreift, während Tiefseeanglerfische mit winzigen leuchtenden Ködern reglos in der Dunkelheit schweben. Knochenweiße Wirbelbögen tauchen auf und verschwinden wieder, wo biolumineszente Impulse vorbeiziehender Tiere ihre Mineraloberflächen streifen, und schwefeldunkle Flecken sowie ein schwacher mikrobieller Glanz überziehen das Sediment rund um die Überreste, Zeichen aktiver Chemosynthese unter der Oberfläche. Weit im Hintergrund, am Rand der fassbaren Dunkelheit, deutet ein rotoranger Schimmer auf einen entfernten hydrothermalen Einfluss hin, während Meeresschnee – Partikel organischen Materials aus den Wasserschichten darüber – lautlos durch diese kalte, druckstarre Welt sinkt und ein Universum beschreibt, das vollständig aus sich selbst heraus existiert.

In einer Tiefe, in der jedes Sonnenlicht seit Hunderten von Metern erloschen ist, treibt ein schwebender Baldachin aus gallertigen Medusen durch stockfinsteres, eiskaltes Wasser von nahezu dreistelligem Atmosphärendruck. Ihre durchscheinenden Schirme und radialen Kanäle entfachen sich in gestaffelten Impulsen aus kaltem Blau-Grün — reines Biolumineszenz, erzeugt durch Photoproteine im lebenden Gewebe —, und formen so eine dreidimensionale, selbstleuchtende Architektur mitten im offenen Pelagial. Nur wo diese Lichtwellen das Wasser streifen, werden vereinzelte Partikel marinen Schnees sichtbar, jene unaufhörliche Rieselung organischer Flocken, die als einziger Nahrungsstrom aus den sonnengeküssten Schichten herabsinkt. Unter dem leuchtenden Schirmdach zieht eine dunklere Silhouette lautlos vorüber — ein Räuber des Mitternachtswassers, dessen Körperbau auf Stealth, Tarnung und den seltenen Augenblick des Überwältigend-Zuschnappenkönnens ausgerichtet ist, genau wie Fangzahnfische, Viperfische oder tintenfischartige Jäger, die in diesen Tiefen seit Jahrmillionen evolviert haben. Diese Welt existiert vollständig in sich selbst: keine Strömung von oben, kein Signal von außen, nur der pulsierende Austausch zwischen Licht und Dunkel, zwischen Treibendem und Fressendem, in einem Ozean, der seine eigenen Gesetze schreibt.

Im Herzen der Mitternachtszone, in Tiefen zwischen tausend und viertausend Metern, bewegt sich ein solitärer Räuber durch die unaufhörlich fallende Partikelwelt des marinen Schnees – winzige Aggregate aus totem organischen Material, Kotpellets und Bakterienflöckchen, die wie ein unendlicher stiller Niederschlag durch schwarzes, nahezu eiskaltes Wasser sinken und zugleich die einzige Verbindung zur fernen, längst erloschenen Oberfläche darstellen. Bei einem Druck von weit über zweihundert Atmosphären und Temperaturen knapp über dem Gefrierpunkt hat die Evolution diesen Körper auf äußerste Effizienz gedichtet: die kastanienbraun-schwarze Pigmentierung löscht jede Reflexion biologischer Biolumineszenz aus der Umgebung aus, während spärlich verteilte Photorhoren an Flanke und Bauch in einem kühlen Cyan kurz aufleuchten und damit entweder Beuteorganismen täuschen oder das Silhouettenrisiko gegen den fernen schwachen Schimmer von oben aufheben – ein Prinzip, das als Gegenlicht-Tarnung bekannt ist. Weit darunter zeichnet sich die abyssale Ebene kaum mehr als eine flüsterfeine Schichtung basaltischen Schlamms ab, akkumuliert über Jahrmillionen, und am äußersten Horizont deutet ein entfernter rotoranger Chemolumineszenz-Schleier auf einen hydrothermalen Plume hin, dessen heiße Fluide sich in Sekunden mit dem umgebenden Tiefenwasser vermischen und eine Nahrungsoase für Bakterien, Invertebrate und all jene schaffen, die in dieser Dunkelheit nach Energie suchen. Diese Welt existiert vollständig in sich selbst: kalt, still, unter unvorstellbarem Druck, und vollkommen gleichgültig gegenüber allem, was außerhalb ihrer schwarzen Wassersäule geschieht.
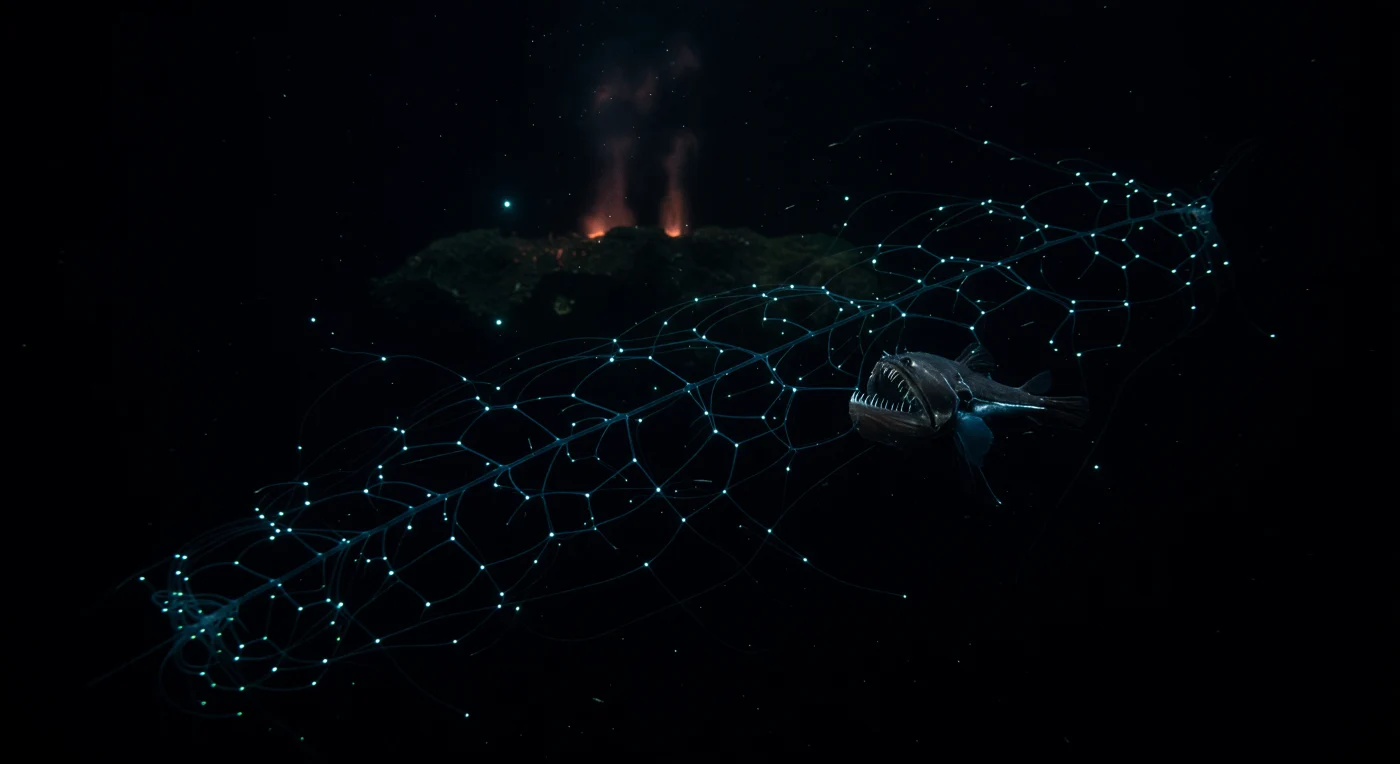
In einer Tiefe zwischen tausend und viertausend Metern, wo kein Sonnenstrahl je ankommt und der hydrostatische Druck mehrere hundert Atmosphären beträgt, entfaltet eine Siphonophore ihr lebendiges Fanggeflecht aus dem schwarzen Wasser — ein koloniales Tier, das genetisch identische Zooide zu einem einzigen, hochspezialisierten Organismus verknüpft, dessen Länge jene der größten Blauwale übertreffen kann. Jeder Knoten dieses diagonalen Gitters aus Tentakeln und Nematocysten sendet ein kühles blau-grünes Biolumineszenzlicht aus, das nicht zur Orientierung dient, sondern zur Lockung: Kleinkrebse und Zooplankton, die in der Wassersäule treiben, werden von den Lichtpunkten angezogen und geraten in die fast unsichtbaren Fangarme. Knapp außerhalb dieser leuchtenden Geometrie verharrt ein Fangzahnfisch (*Chauliodus* sp.) in vollständiger Reglosigkeit — sein gestreckt-schlanker Körper mit metallisch-anthrazitfarbenem Integument ist an das Druckregime angepasst, die überproportionalen Fangzähne fangen nur einen flüchtigen Cyan-Reflex des Siphonophoren-Lichts auf, während er auf eine verwertbare Beute wartet, die sich dem Netz zu sehr nähert. Im fernen Hintergrund steigt ein schwacher orange-roter Schimmer aus hydrothermalen Schloten auf, deren austretende Fluide frisches Basaltgestein prägen und chemosynthetische Gemeinschaften ernähren, während Meeresschnee — organische Partikel, die langsam aus den oberen Wasserschichten herabsinken — die einzige stoffliche Verbindung zur sonnenbestrahlten Welt darstellt, die diese Organismen jemals kennen.

Im Bereich zwischen tausend und viertausend Metern Tiefe, wo kein Sonnenstrahl mehr eindringt und der hydrostatische Druck auf biologische Strukturen bis zu vierhundert Atmosphären ausübt, erhebt sich ein dichter Wald aus Riesenröhrenwürmern (*Riftia pachyptila*) über zerklüftetem Basalt, ihre elfenbeinfarbenen Röhren eng gebündelt, ihre kiemenroten Federkronen vom schwachen chemolumineszenten Schimmer der Hydrothermalquellen beleuchtet. Schwarze Raucher stoßen schwefelreiche Mineralwolken in das tiefblaue Dunkel, während mikrobielle Biofilme und Mineralpartikel das Eigenleuchten der Quellen diffus zerstreuen und feines marines Schneegestöber reglos in kaum spürbaren Strömungen schwebt. An der Peripherie dieser chemosynthetischen Oase wartet ein Anglerfisch (*Melanocetus* sp.) bewegungslos, sein samtiger, druckadaptierter Körper nahezu unsichtbar, die transluzenten Zähne kaum zu erahnen, während das biologische Leuchtorgan über seinem Schädel ein kaltes cyan-grünes Licht pulsiert – eine Evolution der Täuschung, geformt über Jahrmillionen absoluter Dunkelheit. Kleine biolumineszierende Tiere flackern am Kolonierand auf, kurz erhellen sich nasse Chitinpanzer und fleischige Plumosa-Kronen im blauen Blitzen, bevor weiter im Schwarz nur noch andeutungsweise die Nadelsilhouette eines Viperfisches und die gespenstisch langen Tentakelfäden eines *Magnapinna*-Tintenfisches erkennbar werden – Mitglieder einer Raubtiergilde, die ohne jedes Sonnenlicht, in Stille und unter extremem Druck, seit dem Mesozoikum die tiefste Dunkelheit bewohnt.
